«Когда вы наступаете на гвоздь,— говорит Г. Уолтер в своей книге «Живой мозг»,— вы сначала подпрыгиваете и лишь затем ощущаете боль. Сигналы успевают проделать свой путь по рефлекторной дуге туда и обратно за время меньшее, чем требуется чувству боли для достижения мозга».
Не приходится объяснять, что механизмы «подпрыгивания» и боли не одни и те же. Путь болевой реакции гораздо длиннее и во много раз сложнее, чем дуга обычного примитивного рефлекса. Прежде чем достичь высших центров сознания в коре больших полушарий, болевая эстафета проходит через спинной мозг с его сложными входными и выходными устройствами.
Американский физиолог Вулдридж называет спинной мозг главным кабелем организма. И действительно, в его толще проходят миллионы нервных волокон, доставляющих информацию головному мозгу и передающих мышцам и железам инструкции — результаты переработки этой информации нейронами коры и подкорки.
Многочисленные тела нервных клеток, сгруппировавшиеся во внутренней части спинного мозга и образующие похожее на бабочку или на букву Н серое вещество, окружены мощной системой проводящих путей — пучков и канатиков, из которых образуется белое вещество. Анатомы и физиологи выделили в нем восходящие и нисходящие столбы, канатики и пучки, имеющие различное целевое назначение.
Чувствительные нервные корешки вступают в спинной мозг в виде двух более или менее разграниченных пучков. Один пучок, расположенный ближе к средней линии, поднимается кверху и входит в состав задних восходящих столбов спинного мозга. Он состоит из толстых, покрытых миелином волокон, прерывающихся в нейронах продолговатого мозга. По его волокнам в головной мозг передаются импульсы от рецепторов прикосновения и от внутренних органов нашего тела. Эти волокна доходят до зрительных бугров, здесь снова прерываются и затем направляются к коре задней центральной извилины головного мозга.
Волокна второго пучка заканчиваются в задних рогах серого вещества, разветвляясь вокруг нервных клеток. Они передают нейронам импульсы от рецепторов тепла, холода и боли (рис. 9). На верхушках задних рогов находятся скопления мелких клеток, отдаленно напоминающие студень. Это студенистое или желатинозное вещество — желатинозная субстанция Роланда, состоящая из мелких нервных кистей — телец Гирке — Вирхова. Желатинозная субстанция — это замкнутая, высоко специализированная система, простирающаяся вдоль всего спинного мозга, позади задних рогов серого вещества. К ней подходят как толстые, так и тонкие афферентные волокна. В формировании чувства боли она играет, по мнению современных исследователей, особо важную роль. Раздражение ее некоторыми химическими веществами вызывает иногда у животных сильнейшую боль в конечностях и туловище.
Исходя из этого наблюдения, советский ученый Г. Н. Крыжановский разработал интересную модель болевого синдрома у животных. Модель эта была использована и нами для изучения состояния симпато-адреналовой системы при боли. Вот почему мы уделяем ей больше внимания, чем аналогичным работам других авторов.
В область задних рогов крестцово-поясничного отдела спинного мозга животного с помощью тончайшей стеклянной пипетки вводится незначительное количество столбнячного токсина. Не проходит и нескольких часов, как крыса (или кошка) начинает усиленно вылизывать некоторые участки задней конечности на той стороне, где был введен токсин. По всем признакам она испытывает мучительную боль. Постепенно вылизывание делается все более и более ожесточенным, животное набрасывается на больную ногу, кусает ее, кричит, иногда даже выгрызает куски ткани, так что обнажается кость. При этом можно наблюдать все характерные болевые реакции. Интересно, что в пораженной коже резко повышена чувствительность к прикосновениям, уколам, поглаживанию. Едва заметное касание вызывает сильнейшую боль. Животное становится необычайно агрессивным. Описанную модель можно рассматривать как центральный или спинальный болевой синдром.
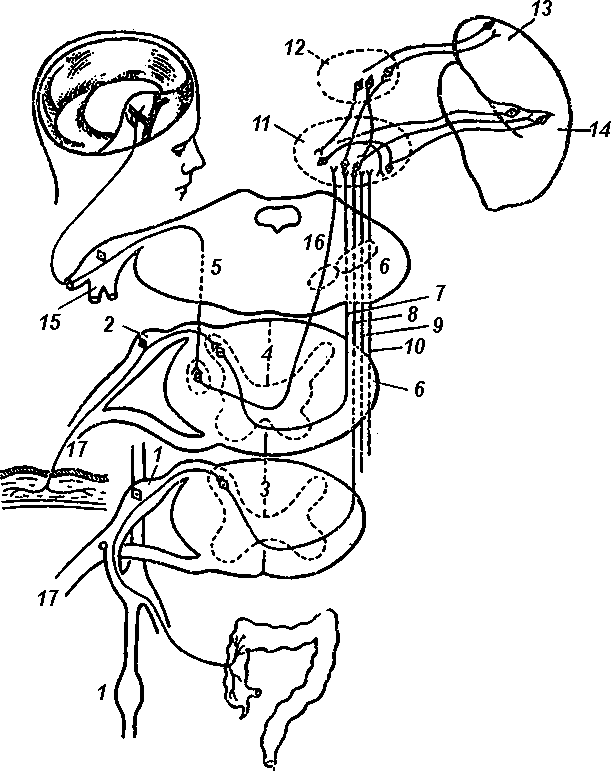
Рис. 9. Пути болевых раздражений (по Гийому, Сеза и Мазарсэ)
1 — симпатический ствол; 2 — межпозвоночный узел; 3 — поперечное сечение спинного мозга на уровне девятого грудного позвонка; 4 — поперечное сечение спинного мозга на уровне первого шейного позвонка, 5 — поперечное сечение на уровне Варолиева моста, 6 — спинно-бугровый тракт; 7 — его шейная часть; 8 — его грудная часть; 9 — его пояснично-крестцовая часть; 10 — его крестцовая часть; 11, 12 — ядра зрительных бугров, 13 — лобная доля мозга; 14 — задняя центральная извилина, 15 — тройничный нерв; 16 — путь от тройничного нерва к зрительному бугру; 17 — путь от болевого рецептора в спинной мозг
От клеток серого вещества начинаются новые волокна, которые, перекрещиваясь, т.е. переходя из правой половины спинного мозга в левую и из левой в правую, объединяются в нервный пучок, известный под названием спинно-бугрового канатика. По нему идут сигналы боли. Этот канатик располагается в передне-боковом столбе спинного мозга. Состоит он в основном из тонких, лишенных миелиновой оболочки волокон. По этому пути болевые и температурные раздражения передаются в большие полушария головного мозга. Конечные нейроны его находятся в теменной доле коры головного мозга. Он является главным коллектором проводящих путей болевых импульсов нашего тела и собирает нервные волокна от многочисленных болевых рецепторов, заложенных во всех органах и тканях организма человека и животных.
Перерезка спинно-бугрового канатика подавляет болевые и температурные ощущения в области, откуда идут перерезанные волокна. При блокаде правого пучка болевые ощущения исчезают в определенных участках левой половины тела, при блокаде левого пучка — в правой.
Однако чувство боли передается не только по спинно-бугровым канатикам. Природа всегда страхует себя и создает резервные возможности. Ей необходим избыток надежности. Это приходится постоянно помнить.
Отдельные болевые и температурные импульсы, не перекрещиваясь и не вступая в главный коллектор болевой чувствительности, направляются в головной мозг через серое вещество задних рогов по так называемым клеткам-передатчикам. Одновременно очень большая часть болевых волокон попадает в спинной мозг более сложным путем. Какой-то отрезок своего длинного пути, начавшегося в периферических рецепторах, они проходят вдоль позвоночного столба в толще пограничной симпатической цепочки. И лишь затем через задние корешки проникают в спинной мозг. Вот почему перерезка спинно-бугрового канатика в некоторых случаях не снимает чувства боли.
Из спинного мозга проводящие волокна без какой-либо заметной границы переходят в продолговатый мозг — важнейший отдел центрального нервного аппарата. Помимо центров дыхания, жевания, глотания, кашля, рвоты, здесь находятся ядра чувствительных нервов черепа и мягких тканей головы — тройничного, языко-глоточного, добавочного и блуждающего. Здесь же находятся промежуточные нейроны, в которых прерываются волокна восходящих путей спинного мозга и начинается вторая дистанция их пути в головной мозг. Неподалеку от спинно-бугрового канатика в том же направлении в толще продолговатого мозга проходят волокна, связывающие ядра черепно-мозговых нервов с высшими болевыми центрами.
Непревзойденное творение природы, ее высшее достижение — головной мозг человека — изучается сотни лет учеными всего мира, но до сих пор мы не можем сказать, что его строение, химический состав и функции полностью известны. Физиологи нередко называют мозг «черным ящиком», используя образное выражение основоположника кибернетики Норберта Винера. Под «черным ящиком» этот выдающийся математик подразумевал устройство, которое выполняет определенную операцию над входным потенциалом, но строение и функции которого, обеспечивающие выполнение операции, нам не известны.
«Можно с правом сказать,— говорит И. П. Павлов[10],— что неудержимый со времен Галилея ход естествознания впервые заметно приостанавливается перед высшим отделом мозга, или, вообще говоря, перед органом сложнейших отношений животных к внешнему миру. И, казалось, что это недаром, что здесь действительно критический момент естествознания, так как мозг, который в высшей его формации — человеческом мозгу — создавал и создает естествознание, сам становится объектом этого естествознания».
Многое изменилось с того времени, когда были произнесены эти слова. С помощью современных методов физического и химического исследования, усилителей электрических токов, световых и электронных микроскопов, спектрографов, микроэлектродов и других сложных приборов, изобретенных гением человека, удалось разрешить многие загадочные и спорные вопросы физиологии головного мозга.
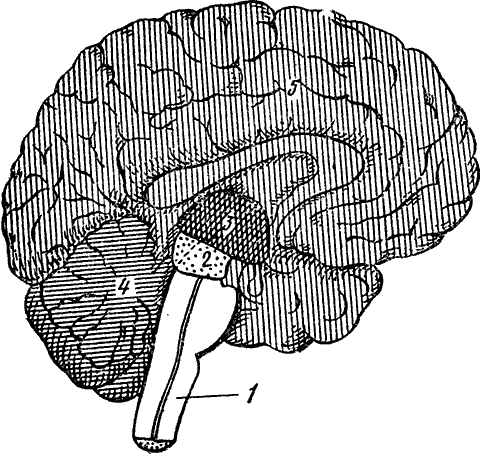
Рис. 10. Продольный разрез мозга (схема)
1 — продолговатый мозг; 2 — средний мозг; 3 — промежуточный мозг; 4 — мозжечок; 5 — передний мозг
Г. Уолтер объясняет успехи в области изучения мозга «приложением техники, фактов и теорий к проблемам человеческого общества в эпоху всеобщей связи, водородных бомб и космических путешествий».
Благодаря этому таинственный орган, перед которым пасовала мысль крупнейших исследователей на всем протяжении истории человечества, постепенно становится доступным познанию во всех своих деталях и своеобразных особенностях. Исследователь начинает постигать сущность процессов, совершающихся в недрах мозга. Понемногу «черный ящик» приоткрывает свои тайны.
Головной мозг составляет передний и наиболее важный отдел центральной нервной системы. Слой серого вещества покрывает полушария головного мозга, образуя его самую совершенную и сложную часть — кору. В толще головного мозга можно обнаружить скопления нервных клеток, образующие так называемые подкорковые центры — высшие, промежуточные и подчиненные, деятельность которых связана с отдельными функциями нашего организма. Густая сеть нервных волокон, объединяющих и связывающих различные центры, пучки выходящих из клеток коры и входящих в нее нервных путей составляют ткань мозга, неповторимую и непревзойденную по своему строению, химическому составу и физиологическому назначению.
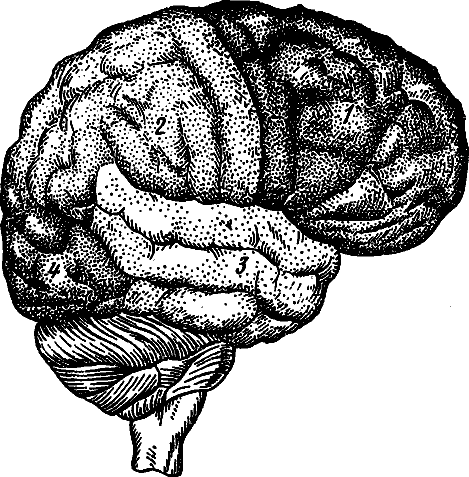
Рис. 11. Наружная поверхность головного мозга (схема)
1 — лобная доля; 2 — теменная доля, 3 — височная доля, 4 — затылочная доля
Головной мозг разделяется на пять отделов: а) передний мозг с полушариями большого мозга, б) промежуточный мозг, в) средний мозг, г) мозжечок, д) продолговатый мозг с Варолиевым мостом (рис. 10). Большой мозг состоит из двух полушарий, на поверхности которых кора образует глубокие борозды и причудливые извилины. Каждое полушарие разделяется на отделы, называемые «долями» — лобной, теменной, затылочной и височной (рис. 11).
Какую же роль играют эти отделы в формировании болевого ощущения? Что делается с болевым сигналом, пробежавшим свой извилистый путь от нервного окончания до высшего распорядительного органа?
Зрительные бугры
Из продолговатого и среднего мозга, миновав Варолиев мост, спинно-бугровый канатик и другие чувствительные проводники вступают в зрительные бугры, наиболее массивные и сложные подкорковые образования больших полушарий головного мозга.
«В физиологии нервной системы,— писал В. М. Бехтерев в 1883 г.,— нет более темной области, как отправление зрительных бугров». И несмотря на то что за истекшие годы появилось немалое число работ, посвященных анатомии и физиологии зрительных бугров (начиная с классических исследований самого Бехтерева, капитальных изысканий Карплюса и Крейдля), роль этой области мозга далеко еще нельзя считать окончательно выясненной.
Лишь в последние годы некоторые стороны деятельности зрительных бугров стали значительно яснее и сделалась более понятной их роль в физиологии больших полушарий мозга.
Зрительные бугры (таламическая область) представляют большие скопления серого вещества в межуточном мозгу. С двух сторон — правой и левой — они образуют стенки третьего желудочка мозга. К их нижней поверхности прилежит подбугорье (гипоталамус), задний утолщенный конец образует так называемую подушку, в которую поступает слуховая и зрительная информация. Огромным числом самых разнообразных нервных путей связаны они буквально со всеми отделами головного и спинного мозга. На разрезе легко обнаружить, что в зрительных буграх имеется пять основных серых ядер, разделенных прослойками белого вещества. Можно считать установленным, что зрительные бугры собирают все чувствительные импульсы, поступающие от периферических рецепторов в центральную нервную систему.
К передней части внутреннего (медиального) ядра подходят волокна обонятельного тракта, к задней — волокна протопатической болевой чувствительности. Здесь, по современным представлениям, формируются чувства «удовольствия» и «неудовольствия». Задняя часть наружного (латерального) ядра является основным центром проприоцептивной чувствительности, т.е. получает импульсы из мышц и суставов, а также тактильные, болевые и температурные сигналы. К ней же подходит болевая информация, поступающая по тройничным нервам. Существование в зрительных буграх специальных «болевых» ядер, клетки которых реагируют только на сверхсильные раздражения, было показано рядом американских и английских исследователей на симпозиуме по боли 1973 г. в Вашингтоне.
Остальные ядра не имеют непосредственного отношения к восприятию боли. Роль их многообразна и зависит в значительной степени от связей с другими отделами мозга. Множество нервных волокон соединяет ядра зрительных бугров с подбугорьем, где заложены центры высших вегетативных функций (обмена веществ, терморегуляции, сосудистого тонуса и т.д.), и с другими подкорковыми образованиями, а также с корой головного мозга. Эти волокна пронизывают всю толщу мозга. Они обеспечивают бесперебойную двустороннюю связь зрительных бугров с корой и коры со зрительными буграми. Зрительные бугры представляют древнейшую часть головного мозга. Они образовались в нервной системе живых существ, населявших Землю за много миллионов лет до появления человека. И лишь впоследствии, в процессе эволюции, на более высоких ступенях зоологической лестницы появилась та часть мозга, которая носит название полушарий.
В течение многих лет шел спор о расположении высших центров болевого ощущения. До недавнего времени считалось наиболее вероятным, что болевое ощущение формируется не в коре головного мозга, а в зрительных буграх. Это предположение, высказанное английскими учеными Гэдом и Холмсом, поддерживали многие виднейшие физиологи и клиницисты (Лейн, Лавастин, Лериш, М. И. Аствацатуров, Л. А. Орбели и др.). Однако большинство исследователей склоняется к мысли, что таламическая область мозга является лишь важнейшим узлом, но все же промежуточным центром болевого восприятия.
В настоящее время физиологи и практические врачи рассматривают зрительные бугры как главный информационный центр головного мозга. К ним направляется поток отдельных раздражений, возникших в различных рецепторах нашего тела. Однако множество импульсов, бомбардирующих эту область мозга, нами не осознается. Они не доходят до коры. Надо полагать, что и не все сверхсильные раздражения, относящиеся к категории алгогенных, т.е. вызывающих боль, воспринимаются и должным образом оцениваются сознанием. Но если человек почувствовал боль, то это значит, что болевые импульсы достигли высшего отдела центральной нервной системы — коры больших полушарий — и осознаны ею как боль.
При некоторых формах нарушения деятельности зрительных бугров, особенно наружного ядра, вызванных недостаточным кровоснабжением, закупоркой артерии, питающей эту область мозга, или, в более редких случаях, опухолью, возникают своеобразные изменения чувствительности, известные в клинике под названием таламического (бугрового) синдрома. Заболевание это, описанное в 1906 г. французскими невропатологами Дежерином и Русси, представляет интереснейший пример центральных болей, не связанных с каким-либо болевым очагом в организме. На Парижском симпозиуме по боли 1967 г. известный французский клиницист Гарсен назвал таламический синдром «болью особого характера». Она ощущается главным образом в одной или нескольких конечностях, в области рта, лица — как правило, на стороне, противоположной очагу поражения.
Бугровые боли могут быть глубокими или поверхностными, но всегда имеют нечеткий, расплывчатый — как говорят врачи, диффузный — характер. Даже самые наблюдательные пациенты не в состоянии дать им точное определение и жалуются обычно на страх, тоску, неожиданные приступы раздражения, ярости, душевной слабости. При этом болевые ощущения отличаются удивительной стойкостью и почти не поддаются лечению.
Лица, страдающие таламическим синдромом, испытывают болезненность в одних участках кожной поверхности и теряют чувствительность в других. Легкое раздражение кожи, прикосновение, толчок вызывают долго длящееся «ползанье мурашек», покалывание и т.д. Эти ощущения сохраняются значительно дольше, чем у здоровых людей. Немецкий невропатолог Ферстер дал этому явлению название гиперпатии.
Возникновение таламических болей разные исследователи объясняют по-разному. Гэд, например, считал, что они возникают в том случае, когда кора головного мозга перестает контролировать деятельность нижележащих отделов центральной нервной системы. Были предложены и другие теории, но они вряд ли интересны читателю-неспециалисту.
В связи с этим возникает очень трудный для теоретического и экспериментального решения вопрос о переработке зрительными буграми получаемой ими информации.
Можно ли считать, что болевая импульсация вообще не доходит до корковых нейронов? Отнюдь нет! И к этому вопросу нам еще придется не раз возвращаться.
Пока же ограничимся некоторыми замечаниями. Мозг человека в состоянии бодрствования получает ежесекундно несметное количество сигналов из внешней и внутренней среды. Если каждый из них дойдет до сознания, жизнь вообще станет невозможной. Для того чтобы мозг осуществлял свою высшую регулирующую и направляющую деятельность, ему необходимо выделять наиболее значимые сигналы из того «шумового фона», который неизбежно возникает в мириадах живых систем, составляющих организм.
Этот отбор сигналов и происходит в низших и промежуточных инстанциях центральной нервной системы.
Если зрительные бугры являются в основном центром древней, грубой, ничем не смягченной (протопатической) чувствительности, то кора головного мозга способна дифференцировать сигналы чувствительности тонкой, призванной смягчить и локализовать чувство боли. Конечные центры эпикритической чувствительности находятся в коре.
Спор между сторонниками «корковой» и «бугровой» локализации болевого восприятия потерял в настоящее время свою остроту.
Вскрылись новые обстоятельства, позволяющие по-новому проследить пути болевых ощущений, совершенно иначе расценить давно известные факты и «устоявшиеся» истины.
Уже давно известно, что при раздражении какого-нибудь чувствующего нерва в коре головного мозга можно обнаружить два вида электрической активности, различающихся по времени их возникновения.
Еще в 1942 г. Морисон и Демпси показали, что и при электрическом раздражении ядер зрительного бугра в коре удается получить два типа электрических реакций. Первый — с коротким латентным периодом, обнаруживающимся преимущественно в чувствительных зонах коры, в корковых ядрах тех или иных анализаторов. Второй — с длинным латентным периодом, распространяющимся по всей коре и не поддающимся точной локализации. Эта распространенная (генерализованная) реакция коры головного мозга наблюдается при раздражении всех видов чувствующих волокон (зрительных, слуховых, обонятельных, тактильных, температурных и болевых). Можно было предположить, что периферические раздражения поступают в кору по двум различным проводящим системам. Один путь — специфический, хорошо известный, знакомый нам во всех деталях. Это путь зрительный, слуховой, вкусовой, обонятельный, тактильный, болевой, проприоцептивной информации. Другой — до недавнего времени неясный, гипотетический, более подробно изученный только за последние годы.
Ретикулярная формация
Хирурги во время операции на центральной нервной системе давно заметили одно весьма интересное обстоятельство. Оказалось, что можно во время операции производить любые разрезы через кору больших полушарий мозга, можно удалять значительные участки мозговой ткани, иногда даже целое полушарие, тем не менее пациент продолжает бодрствовать и не теряет сознания. Однако стоит лишь слегка задеть инструментом некоторые точечные участки в глубине мозга, как больной немедленно впадает в бессознательное состояние, похожее на сон.
В конце 40-х годов нашего столетия два видных физиолога — американец Мэгоун и итальянец Моруцци провели на животных серию экспериментов, которые не только объяснили это наблюдение, но и заложили основу изучения функций особого образования мозга, известного под названием ретикулярной формации.
Ретикулярная формация представляет скопление нервных клеток в центральных отделах той части головного мозга, которую называют мозговым стволом. Нейроны ретикулярной формации благодаря большому числу ветвящихся и переплетающихся отростков образуют густую нервную сеть, откуда и возникло название ретикулярная, или, в переводе на русский язык, сетевидная формация.
Анатомы уже более 100 лет назад описали этот своеобразно построенный отдел мозга. Но физиологическое значение его стало понятным лишь сравнительно недавно.
Если в опытах на животных разрушить определенные участки ретикулярной формации мозгового ствола, сохранив неповрежденными другие мозговые образования, наступает состояние длительной спячки. Вывести животное из спячки практически не удается, применяя даже самые сильные внешние раздражители. Нервные импульсы беспрепятственно достигают коры больших полушарий головного мозга, но животное на них не реагирует. При этом электрические потенциалы, возникающие в коре мозга, ничем не отличаются от потенциалов, наблюдаемых при глубоком естественном сне.
Попробуем через специально вживленные электроды раздражать слабым электрическим током сохранившиеся после первого опыта участки ретикулярной формации. Животное сразу просыпается, а на электроэнцефалограмме появляются типичные сдвиги, называемые физиологами «реакцией пробуждения».
Результаты этих опытов показывают, что ретикулярная формация непосредственно участвует в процессах регулирования сменяющих друг друга состояний — сна и бодрствования. В чем же смысл этого регулирования?
Как известно, мозг человека содержит до 15 млрд. чрезвычайно сложно устроенных нервных клеток. Довольно подробно изучены многочисленные, подчас в высшей степени причудливые связи этих клеток друг с другом. Активное состояние — возбуждение, возникнув в одной клетке, теоретически может распространяться по всему мозгу. Если бы это на самом деле имело место, у нас не было бы никаких оснований восторгаться и изумляться деятельностью мозга. Любое воздействие на организм приводило бы к одной и той же однотипной реакции, напоминающей судорожный припадок, сходный с эпилептическим. А между тем мы прекрасно знаем, как точно приурочены ответы мозга к вызвавшему их воздействию. Достигается это благодаря очень тонкой и четкой системе регулирования физиологических процессов в мозгу. Одни его отделы способны воспринимать и перерабатывать приходящее к ним раздражения, другие лишены этой способности. При возбуждении одних нервных клеток в других, иногда лежащих по соседству, иногда отдаленных, возникает торможение (подавление, угнетение деятельности).
Исследования последних лет показали, что в мозгу имеются особые образования, предназначенные в основном для регулирования состояния и активности его отдельных рабочих частей. Одним из наиболее изученных образований подобного рода и является ретикулярная формация.
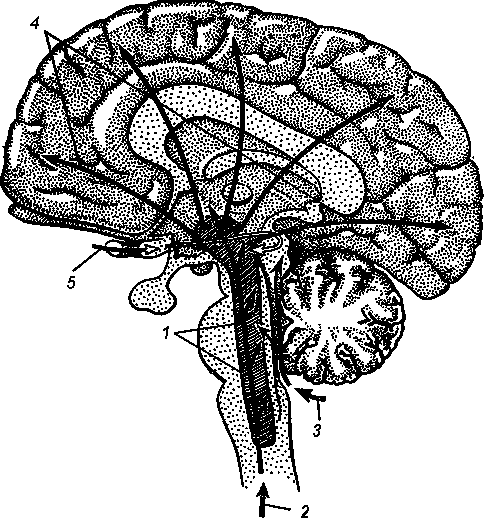
Рис. 12. Активирующие механизмы мозга
1 — ретикулярная формация; 2 — восходящие чувствительные пути спинного мозга; 3 — путь слуховых импульсов; 4 — путь активирующих импульсов к коре головного мозга; 5 — путь зрительных импульсов
Было доказано, что для деятельности головного мозга одного только поступления в кору чувствительных сигналов недостаточно. Необходимо еще определенное рабочее состояние, рабочая настройка клеток коры. И лишь в этом случае могут реализоваться необходимые процессы восприятия и перерабатываться поступившая в кору информация. Такое рабочее состояние элементов коры создает ретикулярная формация. Она оказывает генерализованное, тонизирующее влияние на определенные отделы головного мозга. Внешне это состояние проявляется в виде бодрствования. Вот почему ретикулярная формация получила название активирующей системы мозга (рис. 12). И вот почему книга одного из основоположников учения о ретикулярной формации, Мэгоуна, называется «Бодрствующий мозг».
Наряду с восходящей ретикулярной системой, существует и нисходящая, контролирующая рефлекторную деятельность спинного мозга.
Наличие многочисленных связующих волокон между корой мозга и ретикулярной формацией обеспечивает также обратное регулирующее и направляющее влияние коры на элементы этой системы.
Возникает вопрос, каким же образом ретикулярная формация способна длительно активировать кору больших полушарий, создавать в ней состояние бодрствования?
От всех чувствительных путей, направляющихся в головной мозг и передающих сигналы, вызванные светом, звуком, прикосновением, давлением, теплом, холодом, болью, отходят боковые «дорожки», заканчивающиеся у поверхности клеток ретикулярной формации. Любое внешнее раздражение — вспыхнувшая лампа, стук в дверь, рукопожатие, боль — возбуждает ретикулярную формацию. Она как бы сама «заряжается» энергией и заряжает кору головного мозга (рис. 13).
При этом ответвления от путей, проводящих самую разнообразную информацию, попадают в те же клетки ретикулярной системы. Сигналы, воспринятые ею, теряют свое лицо (или, точнее, свою специфичность). Они становятся лишь инструментом «зарядки», «тонизирования» коры головного мозга. Природа как будто позаботилась о том, чтобы все входные и выходные каналы головного мозга проходили через ретикулярную формацию.
Вначале все казалось очень простым и легко понятным. Ретикулярная формация, благодаря притоку соответствующих импульсов от органов чувств, создает рабочую «атмосферу» в клетках коры и тем самым поддерживает состояние бодрствования. Но на самом деле это — только схема, далеко не исчерпывающая сложных взаимоотношений в центральной нервной системе.
Мы все хорошо знаем, как трудно заснуть после дня, богатого событиями и переживаниями. Если же к этому еще примешиваются тревоги, волнения, заботы, может наступить бессонница, т.е. ненормальное, длительное, как бы неотключаемое состояние бодрствования. Человек не может заснуть, хотя приток раздражений извне даже уменьшен. Хорошо известно, что страдающие бессонницей пытаются выключить даже самые слабые и привычные раздражители. Они останавливают на ночь тикающие часы, занавешивают окна, укутывают голову одеялом.
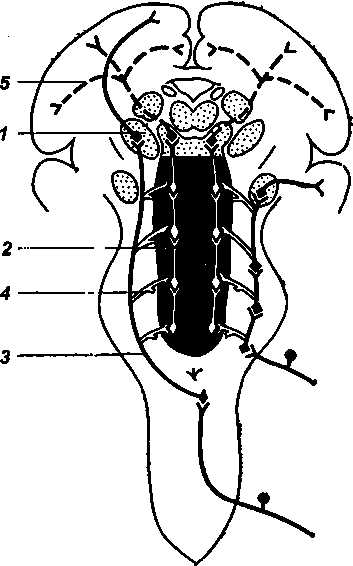
Рис. 13. Схематическое изображение специфических и активирующих систем головного мозга (по Бредли)
1 — ядра зрительных бугров; 2 — ретикулярная формация; 3 — специфический афферентный путь; 4 — ответвления от специфического пути к клеткам ретикулярной формации; 5 — активирующая система
Понять и объяснить эти явления удалось сравнительно недавно, когда было обнаружено, что помимо нервных влияний, передающихся из ретикулярной формации «вверх», к коре больших полушарий, существуют влияния, поступающие в обратном направлении — от коры к ретикулярной формации. Возбуждение, возникшее в определенных участках коры, оказывает в одних случаях стимулирующее, в других подавляющее влияние на ретикулярную формацию. Следовательно, сама кора головного мозга может и поддерживать, и подавлять бодрствование. Наличие системы кольцевых связей (кора — ретикулярная формация — кора) дает возможность высшим отделам центральной нервной системы регулировать через ретикулярную формацию свое рабочее состояние. Этим, вероятно, объясняется то обстоятельство, что лишь некоторые сигналы «заряжают» ретикулярную формацию.
Не все поступающие из внешнего мира раздражения способны прервать наш сон. Спящая мать реагирует на едва заметное движение ребенка, но не просыпается от грохота проносящегося поезда. Многие люди самостоятельно пробуждаются в нужное им время, многие умеют засыпать в любых обстоятельствах, несмотря на свет, шум, разговоры. Кора головного мозга — этот высший распорядитель, распределитель, регулятор деятельности организма — посылает в зависимости от прошлого опыта человека или животного только выборочные, определенные сигналы в ретикулярную формацию.
Можно считать доказанным, что активирующее, «заряжающее», «облегчающее» влияние ретикулярной формации на клетки коры головного мозга — один из факторов, поддерживающих состояние бодрствования. Прекращение сигнализации из ретикулярной формации, блокада ее активирующего влияния вызывает сон. При этом усиливается деятельность «возбуждающих сон» других неспецифических структур мозга, преимущественно расположенных между зрительными буграми и корой. Таким образом, перерыв или ослабление потока восходящих импульсов в кору бодрствующего мозга сопровождается сонливостью, дремотой, сном, потерей сознания. В то же время короткие электрические раздражения ретикулярной формации пробуждают спящее животное. Бодрствование выключается и при некоторых заболеваниях и повреждениях (например при коме, травме черепа). Надо полагать, что это также связано с подавлением активирующих влияний ретикулярной формации на кору мозга.
Однако за последние годы наши представления о механизмах сна и бодрствования значительно расширились и перестроились. Помимо ретикулярной формации, в мозгу были обнаружены и другие структуры, как «пробуждающие», так и «возбуждающие сон». Ушло в область истории и само представление о сне, как о разлитом торможении коры больших полушарий головного мозга. Большинство современных исследователей склоняется к мысли, что во время сна определенные нейроны головного мозга находятся в состоянии повышенной активности, а это приводит к блокаде элементов мозга, получающих информацию из внешней среды. При этом перестраивается вся организация нервных процессов в высших отделах головного мозга.
Сон продолжает оставаться одной из самых трудных загадок физиологии центральной нервной системы. Механизмы его гораздо сложнее, чем это казалось нашим учителям, заложившим основы представлений о сонном торможении, которому мы отдаем треть своей жизни. Старые теории и предположения не выдержали проверки временем, а новые не вышли за границы накопления фактов и недоказанных обобщений. Потребуется еще немало времени и сил, чтобы понять физиологическую и, что еще важнее, физико-химическую сущность сна.
Принято считать, что ретикулярная формация принадлежит к так называемым «неспецифическим» образованиям условного мозга. Ее задача обслуживать специализированные виды его деятельности, в частности анализ сигналов из внешнего и внутреннего мира.
Так ли это на самом деле?
В лаборатории П. К. Анохина был поставлен такой опыт. Кролику наносили болевое раздражение, опуская заднюю лапу в горячую воду, нагретую до 60°. Было отмечено, что при этом в коре головного мозга возникают определенные изменения электрической активности. Если же кролику ввести предварительно аминазин — вещество, блокирующее адренергические элементы ретикулярной формации, болевое ощущение не вызывает сдвигов в электроэнцефалограмме. Да и врачи (особенно психиатры) знают, что аминазин облегчает чувство боли. Но самое интересное заключается в том, что аминазин блокирует не всю ретикулярную формацию, а только те ее элементы, которые возбуждаются под влиянием боли, страха и других отрицательных эмоций. При положительных эмоциональных реакциях, например при кормлении, ретикулярная формация продолжает активировать клетки коры, и животное охотно поедает предложенную ему пищу. Уретан избирательно подавляет реакцию бодрствования, но не затрагивает механизма, активирующего влияние ретикулярной формации на болевое раздражение.
Таким образом, «неспецифическая» ретикулярная формация содержит нервные элементы, влияющие не на всю кору, а только на отдельные комплексы клеток — и, следовательно, обладает специфическим действием на те или другие функции.
Это позволило П. К. Анохину сделать вывод, что активирующая деятельность ретикулярной формации имеет в каждом конкретном случае целенаправленное биологическое значение и вовсе не является «генерализованной», как предполагалось раньше.
В дальнейшем был выявлен еще один интересный факт. Если у крысы разрушить определенные области ретикулярной формации, она начинает поглощать огромные количества пищи, никогда не насыщаясь, и превращается постепенно в жировой ком.
Раздражение током других участков ретикулярной формации вызывает у кошек приступы ярости, а разрушение их превращает послушное, прирученное животное в дикое и агрессивное.
Возникает вопрос, какое же значение имеет такое широко распространенное влияние ретикулярной формации на самые различные проявления деятельности нервной системы, на функции практически всего организма? Каково значение этого влияния, необходимо ли оно, полезно ли?
Вернемся снова в физиологическую лабораторию и проделаем несложный опыт. Через вживленные в различные отделы мозга электроды пропускаем слабый электрический ток в ретикулярную формацию бодрствующего животного. Действие тока скажется почти моментально. Животное «замирает», настораживается, чего-то ждет. Мышцы его напряжены, а на электроэнцефалограмме отмечаются изменения, характерные для высокой активности высших отделов головного мозга.
Такая же картина наблюдается у животного при новом и неожиданном внешнем раздражении, например: свистке, вспышке света, окрике. Подобного рода реакцию И. П. Павлов назвал ориентировочным рефлексом, рефлексом «что?, что такое?», рефлексом на новизну.
Проблема ориентировочной реакции широко разрабатывается и в отечественных, и в зарубежных лабораториях. Исследователи далеко не единодушны в оценке ее сущности и значении.
Что касается высокоорганизованных животных и человека, то правильнее было бы говорить не об ориентировочной, а об ориентировочно-исследовательской реакции со сложными двигательными, вегетативными и поисковыми компонентами.
По-видимому, ориентировочная реакция отражает активную деятельность мозга. Цель ее отобрать среди огромной массы поступающих из внешней среды раздражений лишь те, которые имеют определенное значение для жизнедеятельности и сохранения организма. В осуществлении ориентировочной реакции важную роль играют различные корковые и подкорковые структуры мозга, в том числе ретикулярная формация. Разрушение ее делает ориентировочную реакцию невозможной.
Вот почему ретикулярную формацию можно рассматривать как устройство мозга, обеспечивающее состояние «общей мобилизации» организма по сигналу тревоги. Извне поступил новый и неожиданный сигнал. Пища? Опасность? Друг? Враг? Организм должен быть подготовлен к любой деятельности, которая потребуется: схватить добычу, убежать, спрятаться или не обращать внимания, если тревога окажется ложной. Эта подготовка к любому виду деятельности выражается в переходе высших отделов мозга в более активное состояние, в изменении настройки двигательного аппарата, в обостренном восприятии действительности органами чувств (не поступят ли еще какие-нибудь сигналы, проясняющие обстановку?), в изменении дыхания, работы сердца, сосудов и других внутренних органов. Резервы подтянуты, организм пришел в состояние боевой готовности.
Но вот сигнал повторяется. Теперь уже необходимо произвести анализ обстановки, выработать план действий. На этом этапе мобилизация всех нервных приборов уже не нужна. Требуется лишь настройка тех механизмов нервной системы, которые принимают непосредственное участие в соответствующих неотложных мероприятиях. И высшие отделы головного мозга подавляют деятельность ретикулярной формации. Приказ об общей мобилизации отменен. Ориентировочный рефлекс угасает...
Работами многочисленных советских и зарубежных авторов установлено, что клетки ретикулярной формации возбуждаются не только физическим сигналом. Они чутко реагируют на меняющееся содержание в крови углекислоты, сахара, некоторых химических веществ — и в первую очередь гормонов и медиаторов (адреналина, норадреналина, ацетилхолина, серотонина, гистамина). Так, например, при голодании, когда «голодная» кровь омывает ретикулярную формацию, она мобилизует нервные механизмы поведения, направленные на поиски и добывание пищи. Как остроумно отметил один ученый, она «способствует переводу внутренних потребностей организма в поведение». Затрудненное дыхание у спящего человека приводит к накоплению углекислоты в крови, что вызывает возбуждение ретикулярной формации и пробуждение. Это спасает человека от удушения во сне. Французский ученый Бессон обнаружил, что некоторые клетки ретикулярной формации возбуждаются брадикинином. Введение этого препарата в артерии, снабжающие кровью определенные участки ретикулярной формации, вызывает резкое болевое ощущение.
Еще в начале нашего столетия И. П. Павлов, изучая образование условных рефлексов у животных, подчеркивал особое значение для психической деятельности подкорковых образований мозга, «заряжающих» энергией кору больших полушарий. Он говорил о «слепой силе» подкорки, о подкорке как «источнике энергии» для коры. И на самом деле, теперь уж ни у кого не возникает сомнений, что подкорка является своеобразным аккумулятором энергии и в ней находится особое регулирующее «зарядное» устройство — ретикулярная формация. Разумеется, она не представляет единственный регулирующий механизм мозга. Природа слишком изобретательна и предусмотрительна. В мозгу имеются и другие регулирующие аппараты, и любые попытки некоторых зарубежных исследователей поставить ретикулярную формацию во главе всей деятельности мозга должны быть отвергнуты как не соответствующие научной истине.
Изучение функций ретикулярной формации головного мозга явилось важным шагом вперед на пути познания конкретных материальных механизмов головного мозга. Путь этот дальний, трудный и неровный, но некоторую, небольшую его часть мы прошли.
Какое же отношение имеет ретикулярная формация к восприятию боли?
Можно считать установленным, что при любом раздражении поверхности кожи поступающие с периферии импульсы заряжают ретикулярную формацию мозгового ствола.
Болевое раздражение, пройдя длинный путь от рецептора до головного мозга, воспринимается клетками чувствительной зоны коры. Но вслед за этим через какой-то короткий промежуток времени, исчисляемый тысячной долей секунды, наступает широкая активация коры, обусловленная восходящими, облегчающими влияниями ретикулярной формации.
Существуют два механизма возникновения чувства боли в головном мозгу, связанные у здорового человека воедино. При раздражении одного механизма (специфического) возникает простое ощущение боли; при раздражении второго механизма (неспецифического) — сложное, комплексное ощущение, обозначаемое как страдание, недуг, болезнь. Это ощущение осуществляется при участии целого ряда корковых и подкорковых образований. Ретикулярная формация мобилизует кору для реакции на болевое раздражение, так что последняя оказывается в состоянии привести в действие многообразную систему мероприятий, необходимых для устранения источника боли или для компенсации вызываемых ею нарушений жизнедеятельности организма.
Можно считать доказанным, что во время болевого раздражения ретикулярная формация посылает в кору головного мозга огромное число нервных сигналов, это приводит к резкому изменению активности корковых нейронов. Такие же сигналы идут в кору из зрительных бугров и других подкорковых образований головного мозга.
Кора больших полушарий мозга
Кора больших полушарий связана нервными путями со всеми нижележащими отделами центральной нервной системы, а через них — со всеми органами тела. С одной стороны, импульсы, поступающие с периферии, доходят до той или иной точки коры, с другой — кора посылает «распоряжения» в нижележащие отделы мозга, а оттуда — к различным органам.
Кора головного мозга осуществляет тончайшее равновесие между организмом и внешней средой, регулирует и направляет физиологические процессы, протекающие внутри организма, обеспечивает его сложнейшее функциональное единство.
Каждый анализатор (например, зрения, обоняния, слуха и т.д.) имеет, по представлению И. П. Павлова, в коре головного мозга центральную часть (ядро), где осуществляется высший анализ и синтез, и широкую периферическую зону, в которой аналитические и синтетические процессы совершаются в элементарном виде. Между ядрами отдельных анализаторов разбросаны и перемешаны нервные элементы, принадлежащие различным анализаторам. Если ядро анализатора в силу каких-либо причин разрушено или выбыло из строя, его функцию перенимают периферические элементы того же анализатора. Современная физиология отвергает и узкий («абсолютный») локализационизм и принцип однородности, равноценности всех участков коры мозга. Локализация существует, но имеет «подвижный», «динамический» характер, о чем еще много лет назад говорил И. П. Павлов.
Нервные образования, которые мы привыкли называть «центрами», не ограничиваются корой головного мозга. Они включают и подкорковые структуры, значение которых необычайно велико.
Следует помнить, что любой центр коры головного мозга теснейшим образом связан со всеми другими отделами центральной нервной системы. В этом объединении, или, как говорят, интеграции, и заключается ведущая роль коры мозга в организме.
Представление о единых корковых центрах, полностью обеспечивающих какую-либо определенную функцию, является в настоящее время пройденным этапом в физиологии.
К тому же кора головного мозга отличается необычайной пластичностью, и одни отделы ее легко перенимают функции других, компенсируя расстройство их деятельности, вызванное различными причинами. Наиболее важная задача современной науки выявить анатомическую основу физиологических процессов и одновременно установить связи и взаимосвязи между всеми явлениями, наблюдаемыми в головном мозгу.
Исследования, проведенные различными авторами, как отечественными, так и зарубежными, показали, что в центральной извилине мозга, расположенной спереди от центральной борозды, находится специальная двигательная область. Раздражение ее электрическим током вызывает сокращение определенных мышц противоположной стороны тела. Напротив, удаление этой области хирургическим путем ведет к расстройству координированных движений, шаткости походки, ослаблению мышц. У человека ранение двигательной области сопровождается обычно параличами и другими тяжелыми нарушениями деятельности организма.
С помощью метода условных рефлексов удалось показать, что так называемые двигательные центры содержат чувствительные клетки, к которым приходят периферические раздражения от двигательного аппарата (костей, суставов, мышц). Эта область является мозговым концом двигательного анализатора в такой же степени, как затылочная — мозговым концом зрительного анализатора, височная — слухового анализатора и т.д. В двигательной области имеются как чувствительные клетки, расположенные в верхних слоях коры, так и двигательные, сосредоточенные в ее нижних слоях. Импульсы от рецепторов двигательного аппарата поступают в чувствительные клетки передней мозговой извилины, а отсюда уже передаются двигательным клеткам головного и спинного мозга.
Таким образом, каждый двигательный акт, каждое так называемое произвольное, волевое движение детерминировано, обусловлено раздражениями, поступающими в кору головного мозга из внешней или внутренней среды.
Позади центральной борозды расположена чувствительная область коры. В ней заканчивается путь, начавшийся в рецепторах кожи и внутренних органов. Здесь расположен его конечный этап. Каждое полушарие мозга связано в основном с противоположной половиной тела. Однако существуют связи полушария и с одноименной половиной тела.
Разрушение задней центральной извилины вызывает нарушение чувствительности в соответствующих сегментах тела.
Еще не так давно принято было считать, что рецепторы внутренних органов не имеют своего представительства в этих отделах мозга. Однако работы последних лет показали, что и интерорецепторы имеют связь с корой, хотя раздражение их не вызывает определенных ощущений и не доходит до сознания.
В этом плане особого внимания заслуживают исследования В. Н. Черниговского, его учеников и сотрудников. Исследуя метод отведения электрических потенциалов от определенных участков коры мозга, они показали, что все внутренние органы, посылающие информацию в головной нерв по чревным и брыжеечным нервам, имеют свое представительство в задней мозговой извилине коры мозга. Даже блуждающие нервы — эти мощные проводники импульсов из внутренних органов в центральную нервную систему — тесно связаны с нейронами коры. Открытие это тем более важное и неожиданное, что ядра блуждающих нервов в продолговатом мозгу давно уже описаны и изучены.
Каждый по личному опыту знает, что психические волнения, переживания, неожиданные известия, напряженная умственная деятельность сопровождаются изменениями со стороны сердечно-сосудистой системы, желудочно-кишечного тракта, мочевого пузыря и т.д. У лиц, страдающих грудной жабой или язвенной болезнью желудка, приступы болей возникают обычно при всякого рода психических травмах, волнениях, огорчениях и т.д.
Раздражая электрическим током некоторые области коры, выключая их хирургическими или фармакологическими методами, исследователи установили, что можно по желанию изменять, ослаблять, усиливать, перестраивать работу сердца, желудка, кишок, почек и других внутренних органов. Бесспорно действие коры головного мозга на мочевой пузырь и прямую кишку, доказано ее влияние на слюно- и потоотделение, на состояние мышц, на просвет сосудов и т.д. Условные рефлексы образуются не только при действии какого-либо внешнего агента, но и при всяком раздражении интерорецепторов. Условным раздражением может служить импульс со слизистой оболочки желудка, кишечника, мочевого пузыря, с рецепторов сосудов, селезенки и т.д. Любой сигнал из внутренних органов, если его сочетать с безусловным раздражением, может стать условным стимулом оборонительной пищевой или какой-либо другой деятельности животного.
Это говорит о том, что импульсы из внутренних органов обязательно проникают в кору головного мозга, так как только в этом случае возможна выработка на них условных рефлексов.
Однако почему же все-таки сознание здорового, нормального человека не воспринимает информацию, поступающую из внутренних органов?
В. Н. Черниговский выдвигает три предположения, каждое из которых требует, по-видимому, экспериментальной проверки. Во-первых, общая площадь коры мозга, занимаемая представительством органов чувств, намного больше площади, в которую поступает сигнализация из внутренних органов. Число нейронов, «обслуживающих» в коре рецепторы слуха, зрения, вкуса, обоняния, осязания, во много раз превышает число их, связанных с интерорецепторами. Во-вторых, информация от внутренних органов, в том числе и болевая, идет в мозг по самому медленному пути (по волокнам С), а от других рецепторов — по волокнам А. Сигналы от кожи, мышц, органов чувств гораздо быстрее достигают коры и, возможно, блокируют корковые элементы сознания. В-третьих, возможно, что организм каким-то образом тормозит, задерживает поступление информации из внутренних органов в кору. Этим он спасает наше сознание от огромного потока импульсов, которые могли бы подавить своей силой и количеством деятельность больших полушарий. Быть может, в первую очередь это относится к шестому чувству — к боли, которая должна быть «осознана» корой только при исключительных обстоятельствах.
В своих последних работах В. Н. Черниговский высказывает еще более интересную мысль. «Кора больших полушарий,— говорит он,— действительно может управлять любыми процессами в организме (а значит — и регулировать их). Но она делает это не в порядке обычной работы, а лишь при определенных условиях». И дальше, «...кора больших полушарий в естественных условиях не использует поистине безграничных возможностей; она вмешивается только тогда, когда в этом есть физиологическая необходимость, а также когда экспериментатор нарочито создает соответствующую ситуацию» [11].
В верхней части задней центральной извилины мозга располагаются центры, воспринимающие ощущения из нижних конечностей, в нижней части — центры рецепторов кожной поверхности головы, лица и шеи.
В коре затылочных долей обоих полушарий находятся зрительные центры, в височной доле — слуховые.
Если вырезать у собаки затылочные и височные доли больших полушарий, у нее начинают проявляться некоторые отклонения со стороны зрения и слуха от нормальной их деятельности. После удаления затылочных долей собака не теряет способности видеть. Она обходит встречающиеся на пути предметы, различает свет и темноту — и в то же время не узнает хозяина. У нее разрушен мозговой конец зрительного анализатора, и это лишает ее возможности производить тонкий анализ зрительных раздражений. То же самое происходит и при разрушении мозгового конца звукового анализатора. Животное отличает тишину от звука, но совершенно не в состоянии дифференцировать звуки, разобраться в шумах, тонах и т.д.
Иногда у людей наблюдаются заболевания, называемые «психической слепотой» или «психической глухотой». Такие люди хорошо видят предметы, но не в состоянии их узнать. Они хорошо, слышат, но не понимают слов, не воспринимают речи и музыки.
Эти заболевания возникают в тех случаях, когда структура коры головного мозга в затылочной или височной области нарушена. Нервные окончания у них в порядке, не пострадали и проводящие пути. Импульсы исправно передаются в головной мозг, но разрушено корковое ядро зрительного или слухового анализатора и, следовательно, расстроена аналитическая деятельность коры головного мозга. Большое внимание уделяют в настоящее время исследователи корковой локализации памяти, эмоций, мышления, даже характера, но в нашу задачу не входит подробный разбор бесконечного количества работ, посвященных представительству различных физиологических функций в коре больших полушарий. Ограничимся только вопросами боли.
Задняя центральная извилина воспринимает, по-видимому, также и болевое ощущение. Правда, вопрос этот нельзя считать окончательно решенным. Мнения исследователей разошлись, и если одним удавалось вызвать чувство боли при раздражении электрическим током задней центральной извилины, то другие повторить этого опыта не сумели.
Еще в 1883 г. в клинике профессора И. П. Мержеевского В. М. Бехтерев наблюдал значительное ослабление болевой чувствительности на противоположной половине тела, удаляя у собак участки коры мозга в верхней части височной области.
С тех пор накопилась большая, но весьма разноречивая литература по вопросу о локализации болевых центров в коре головного мозга. Хорошо известны опыты немецкого невролога Ферстера, который, раздражая электрическим током кору головного мозга в области задней центральной извилины, не мог вызвать отчетливого ощущения боли. Однако, как было установлено более поздними исследованиями, раздражение верхней теменной доли, особенно внутренней ее поверхности, сопровождается обычно сильной болью. Некоторые авторы наблюдали болевую реакцию при раздражении электрическим током отдельных участков коры головного мозга и мозжечка.
По-видимому, при таком воздействии усиливаются только уже имеющиеся боли, особенно во внутренних органах, вызванные тем или другим болезненным процессом. Таково, по крайней мере, мнение Суита — одного из видных американских ученых.
Советский физиолог Д. М. Гедеванишвили обнаружил в затылочной доле головного мозга участок коры, раздражение которого слабым электрическим током вызывает у кошек сильнейшую «реакцию гнева». Кошки становятся необычайно агрессивными, начинают мяукать и кричать. У них появляются характерные для боли движения хвоста, приходят в движение брови и усы. Животные кричат, царапаются, рвутся из рук экспериментатора, выпускают когти и т.д.
Наблюдаемая картина, по мнению автора, воспроизводит болевую реакцию у кошки, вызванную раздражением периферического чувствительного нерва. Исходя из этого, Гедеванишвили пришел к выводу, что «реакция гнева» зависит от возбуждения специального центра болевого ощущения. Дальнейшие его опыты показали, что раздражение «центра боли» приводит к повышению кровяного давления, сердцебиению, одышке и т.п.
Все эти опыты требуют проверки и уточнения. Вряд ли в коре головного мозга имеется строго локализованный «центр боли». Надо полагать, что наблюдаемая реакция вызывается не болью, а возбуждением корковых вегетативных центров. Большинство исследователей склоняется к мысли, что у человека болевая чувствительность связана с теменной долей головного мозга и задней центральной извилиной. Однако аффективную, эмоциональную окраску чувство боли приобретает под влиянием лобных долей головного мозга. Одно время при лечении некоторых душевных заболеваний производилась перерезка нервных путей, связывающих лобные доли с другими частями мозга, так называемая лобо- или лейкотомия. В этих случаях чувство боли не исчезало, но боль становилась безразличной, как бы далекой от реального мира.
Вулдридж рассказывает об одном из таких случаев. Врач, беседуя с больной, подвергшейся операции лоботомии, был несколько удивлен, когда больная заявила, что боли у нее не только не исчезли, но даже ничуть не уменьшились. Между тем она выглядела бодрой, спокойной, здоровой и совершенно перестала жаловаться на мучившие ее до операции ощущения. В дальнейшем выяснилось, что лоботомия привела не к ослаблению самой боли, а к такому изменению психического состояния больной, в результате которого боль перестала ее беспокоить. Таких наблюдений накопилось немалое число, но это еще не значит, что для смягчения болей следует прибегать к столь радикальному воздействию.
Сходные результаты были получены при электрическом раздражении некоторых участков ретикулярной формации головного мозга через вживленные в нее электроды. Боли прекратились, но по прекращению раздражения возобновлялись с прежней силой.
В настоящее время считается общепринятым, что раздражение специфических нервных окончаний кожи, мышц, внутренних органов воспринимается как болевое ощущение корковыми клетками головного мозга.
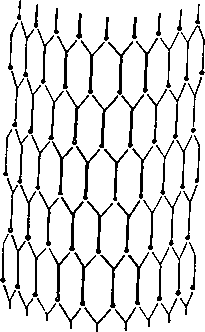
Рис. 14. Схема полисинаптической передачи импульса в сером веществе спинного мозга (по Норденбосу)
Кора больших полушарий играет основную роль в восприятии и «осознании» боли. Субъективное ощущение боли формируется именно в коре. Здесь объединяются и связываются в единое целое все раздражения, поступающие с периферии по разнообразным центростремительным путям. Именно в коре происходит оценка раздражения, сопоставление его с предшествующим опытом, принимается решение и диктуется действие. Частично эти пути заканчиваются в зрительных буграх, но благодаря наличию богатой сети корково-бугровых волокон, благодаря ретикулярной формации так или иначе связываются с клетками высших отделов мозга.
Таким образом, болевой путь от рецептора до коры мозга многократно прерывается в своеобразных реле — нервных клетках, дающих начало новой болевой дистанции. Его можно назвать многодистанционным, или, вернее, много- (поли) синаптическим, т.е. состоящим из цепи, связанных через синапсы друг с другом нейронов и их аксонов. На рис. 14 представлена схема, изображающая нервную афферентную сеть в сером веществе спинного мозга в представлении голландского хирурга Норденбоса. Нервное возбуждение, возникшее в одной точке этого разветвленного дерева, распространяется по огромному числу нейронов (они изображены в виде точек) и, переходя с одного участка на другой, доходит до высших нервных центров.
Осознание и дифференцировка болевого ощущения происходит в высшем распорядительном отделе центральной нервной системы — в коре больших полушарий при участии целого ряда подкорковых образований головного мозга.